H-Evolution-007Animals
Evolution
of Animals
Origin
of Animal Diversity
Animal life began in Precambrian seas with
the evolution of multiceluar creatures that ate other organisms.
We are among their descendants.
What
is an Animal?
Animals are
eukaryotic, multicellular, heterotrophic organisms that obtain nutrients by
ingestion -- ingestion means eating food. This mode of nutrition contrasts
animals with fungi, which obtain nutrients by absorption after digesting the
food outside the body. Animals digest their food within their bodies after
ingesting other organisms, dead or alive, whole or by the piece (Figure 17.2).
Most animals
reproduce sexually. The zygote (fertilized egg) develops into an early
embryonic stage called a blastula, which is usually hollow ball of cells
(Figure 17.3). The next embryonic stage in most animals is a gastrula, which
has layers of cells that will eventually form the adult body parts. The
gastrula also has a primitive gut, which will develop into the animal's
digestive compartment. Continued development, growth, and maturation transform
some animals directly from the embryo into an adult. However many animals
include larval stages. A larva is a sexually immature form of an animal. It is
anatomically distinct from the adult form, usually eats different foods, and
may even have a different habitat. Think how different a frog is from its
larval form, which we calla tadpole. A change of body form, called
metamorphosis, eventually remodels the larva into the adult form.


Figure 17.2 Nutrition by ingestion, the animal way of life. Most
animals ingest relatively large pieces of food, though rarely as large as the
prey in this case. In this amazing scene, a rock python is beginning to ingest
a gazelle. The snake will spend two weeks or more in a quiet place digesting
its meal.
Figure 17.3 Life cycle of a sea star as an example of animal
development. (1) Male and female adult animals produce haploid gametes (eggs
and sperm) by meiosis. (2) An egg and a sperm fuse to produce a diploid zygote.
(3) Early mitotic divisions lead to an embryonic stage called a blastula,
common to all animals. Typically, the blastula consists of a ball of cells
surrounding a hollow cavity. (4) Later, in the sea star and many other animals,
one side of the blastula cups inward, forming an embryonic stage called a
gastrula. (5) The gastrula develops into a saclike embryo with a two-layered
wall and an opening at one end. Eventually, the outer layer (ectoderm) develops
into the animal's epidermis (skin) and nervous system. The inner layer
(endoderm) forms the digestive tract. Still later in development, in most
animals, a third layer (mesoderm) forms between the other two and develops into
most of the other internal organs (not shown in the figure). (6) Following the
gastrula, many animals continue to develop and then mature directly into
adults. But others, including the sea star, develop into one or more larval
stages first. (7) The larva undergoes a major change of body form, called
metamorphosis, in becoming an adult-a mature animal capable of reproducing
sexually.
Most animals have
muscle cells, as well as nerve cells that control the muscles. The evolution of
this equipment for coordinated movement enhanced feeding, even enabling some
animals to search for or chase their food. The most complex animals, of course,
can use their muscular and nervous systems for many functions other than
eating. Some species even use massive networks of nerve cells called brains to
think.

Figure 17.4 One hypothesis for a sequence of stages in the origin
of animals from a colonial protist. (1) The earliest colonies may have
consisted of only a few cells, all of which were flagellated and basically
identical. (2) Some of the later colonies may have been hollow spheres-floating
aggregates of heterotrophic cells-that ingested organic nutrients from the
water. (3) Eventually, cells in the colony may have specialized, with some
cells adapted for reproduction and others for somatic (non-reproductive)
functions, such as locomotion and feeding. (4) A simple multicellular organism
with cell layers may have evolved from a hollow colony, with cells on one side
of the colony cupping inward, the way they do in the gastrula of an animal
embryo (see Figure 17.3). (5) A layered body plan would have enabled further
division of labor among the cells. The outer flagellated cells would have
provided locomotion and some protection, while the inner cells could have
specialized in reproduction or feeding. With its specialized cells and a simple
digestive compartment, the proto-animal shown here could have fed on organic
matter on the sea floor.

Figure 17.5 A Cambrian seascape. drawing based on fossils
collected at a site called Burgess Shale in British Columbia, Canada.
Early Animals and the Cambrian Explosion
Animals probably evolved from a colonial,
flagellated protist that lived in Precambrian seas (Figure 17.4) .By the late
Precambrian, about 600-700 million years ago, a diversity of animals had
already evolved. Then came the Cambrian explosion. At the beginning of the
Cambrian period, 545 million years ago, animals underwent a relatively rapid
diversification. During a span of only about 10 million years, all the major
animal body plans we see today evolved. It is an evolutionary episode so
boldly marked in the fossil record that geologists use the dawn of the Cambrian
period as the beginning of the Paleozoic era. Many of the Cambrian animals seem
bizarre compared to the versions we see today, but most zoologists now agree
that the Cambrian fossils can be classified as ancient representatives of
contemporary animal phyla (Figure 17.5).
What ignited the Cambrian explosion?
Hypotheses abound. Most researchers now believe that the Cambrian explosion
simply extended animal diversification that was already well under way during
the late Precambrian. But what caused the radiation of animal forms to
accelerate so dramatically during the early Cambrian? One hypothesis emphasizes
increasingly complex predator-prey relationships that led to diverse
adaptations for feeding, motility, and protection. This would help explain why
most Cambrian animals had shells or hard outer skeletons, in contrast to
Precambrian animals, which were mostly soft-bodied. Another hypothesis focuses
on the evolution of genes that control the development of animal form, such as
the placement of body parts in embryos. At least some of these genes are common
to diverse animal phyla. However, variation in how, when, and where these genes
are expressed in an embryo can produce some of the major differences in body
form that distinguish the phyla. Perhaps this developmental plasticity was
partly responsible for the relatively rapid diversification of animals during
the early Cambrian. In the last half
billion years, animal evolution has mainly generated new variations of old
"designs" that originated in the Cambrian seas.

Animal
Phylogeny
Because animals
diversified so rapidly on the scale of geologic time, it is difficult, using
only the fossil record, to sort out the sequence of branching in animal
phylogeny. To reconstruct the evolutionary history of animal phyla, researchers
must depend mainly on clues from comparative anatomy and embryology. Molecular
methods are now providing additional tools for testing hypotheses about animal
phylogeny. Figure 17.6 represents one set of hypotheses about the evolutionary
relationships among nine major animal
phyla. The circled numbers on the tree highlight four key evolutionary branch
points, and these numbers are keyed to the following discussion.
(1) The first branch point distinguishes sponges from all other
animals based on structural complexity. Sponges, though multicellular, lack the
true tissues, such as epithelial (skin) tissue, that characterize more complex
animals.
(2) The second major
evolutionary split is based partly on body symmetry: radial versus bilateral.
To understand this difference, imagine a pail and shovel. The pail has radial
symmetry. identical all around a central axis. The shovel has bilateral
symmetry, which means there's only one way to split it into two equal
halves-right down the midline. Figure 17.7 contrasts a radial animal with a
bilateral one.
The symmetry of
an animal generally fits its lifestyle. Many radial animals are sessile forms
(attached to a substratum) or plankton (drifting or weakly swimming aquatic
forms). Their symmetry equips them to meet the environment equally well from
all sides. Most animals that move actively from place to place are bilateral. A
bilateral animal has a definite "head end" that encounters food,
danger, and other stimuli first when the animal is traveling. In most bilateral
animals, a nerve center in the form of a brain is at the head end, near a
concentration of sense organs such as eyes. Thus, bilateral symmetry is an
adaptation for movement, such as crawling, burrowing, or swimming.
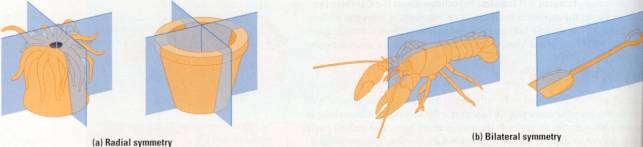
Figure 17.7 Body symmetry. (a) The parts of a radial animal, such
as this sea anemone, radiate from the center. Any imaginary slice through the
central axis would divide the animal into mirror images. (b) A bilateral
animal, such as this lobster, has a left and right side. Only one imaginary cut
would divide the animal into mirror-image halves.
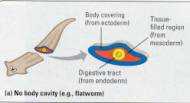
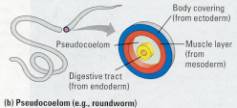
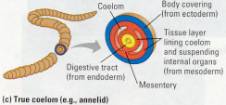
Figure 17.8 Body plans of bilateral
animals. The various organ systems of these animals develop from the three tissue
layers that form in the embryo. (a) Flatworms are examples of animals that lack
a body cavity. (b) Roundworms have a pseudocoelom, a body cavity only partially
lined by mesoderm, the middle tissue layer. (c) Earthworms and other annelids
are examples of animals with a true coelom. Acoelom is a body cavity completely
lined by mesoderm. Mesenteries, also derived from mesoderm, suspend the organs
in the fluid-filled coelom.
(3) The evolution of body
cavities led to more complex animals. A body cavity is a fluid-filled space
separating the digestive tract from the outer body wall. A body cavity has many
functions. Its fluid cushions the suspended organs, helping to prevent internal
injury. The cavity also enables the internal organs to grow and move independently
of the outer body wall. If it were not for your body cavity, exercise would be
very hard on your internal organs. And every beat of your heart or ripple of
your intestine would deform your body surface. It would be a scary sight. In
soft-bodied animals such as earth worms, the noncompressible fluid of the body
cavity is under pressure and functions as a hydrostatic skeleton against which
muscles can work. In fact, body cavities may have first evolved as adaptations
for burrowing.
Among
animals with a body cavity, there are differences in how the cavity develops.
In all cases, the cavity is at least partly lined by a middle layer of tissue,
called mesoderm, which develops between the inner ( endoderm) and outer (
ectoderm) layers of the gastrula embryo. If the body cavity is not completely
lined by tissue derived from mesoderm, it is termed a pseudocoelom (Figure
17.8) .A true coelom. the type of body cavity humans and many other animals
have, is completely lined by tissue derived from mesoderm.
(4)
Among animals with a true coelom. there are two main evolutionary
branches. They differ in several details of embryonic development. Including
the mechanism of coelom formation. One branch includes mollusks (such as clams,
snails, and squids). annelids (such as earthworms), and arthropods (such as
crustaceans, spiders. and insects). The two major phyla of the other branch are
echInoderms (such as sea stars and sea urchIns) and chordates (including humans
and other vertebrates).
Major Invertebrate Phyla
Living as we do on land, our sense of
animal diversity is biased in favor of vertebrates, which are animals with a
backbone. Vertebrates are well represented on land in the form of such animals
as reptiles, birds, and mammals. However, vertebrates make up only one
subphylum within the phylum Chordata, or less than 5% of all animal species. If
we were to sample the animals in an aquatic habitat, such as a pond, tide pool,
or coral reef, we would find ourselves in the realm of invertebrates. These are
the animals without backbones. It is traditional to divide the animal kingdom
into vertebrates and invertebrates, but this makes about as much zoological
sense as sorting animals into flatworms and nonflatworms. We give special
attention to the vertebrates only because we humans are among the backboned
ones. However, by exploring the other 95% of the animal kingdom-the
invertebrates-we'll discover an astonishing diversity of beautiful creatures
that too often escape our notice.
Sponges
Sponges (phylum
Porifera) are sessile animals that appear so sedate to the human eye that the
ancient Greeks believed them to be plants (Figure 17.9). The simplest of all
animals, sponges probably evolved very early from colonial protists. Sponges
range in height from about 1 cm to 2 m. Sponges have no nerves or muscles, but
the individual cells can sense and react to changes in the environment. The
cell layers of sponges are loose federations of cells, not really tissues,
because the cells are relatively unspecialized. Of the 9,000 or so species of
sponges, only about 100 live in fresh water; the rest are marine.


Figure 17.9 Sponge
Figure 17.10 Anatomy of a sponge. Feeding cells called choanocytes
have flagella that sweep water through the sponge's body. Choanocytes trap
bacteria and other food particles, and amoebocytes distribute the food to other
cells. To obtain enough food to grow by 100 g (about 3 ounces), a sponge must
filter 1,000 kg (about 275 gallons) of seawater.
The body of a
sponge resembles a sac perforated with holes (the name of the sponge phylum,
Porifera, means "pore bearer"). Water is drawn through the pores into
a central cavity, then flows out of the sponge through a larger opening (Figure
17.10). Most sponges feed by collecting bacteria from the water that streams
through their porous bodies. Flagellated cells called choanocytes trap bacteria
in mucus and then engulf the food by phagocytosis. Cells called arnoebocytes
pick up food from the choanocytes, digest it, and carry the nutrients to other
cells. Amoebocytes are the "do-all" cells of sponges. Moving about by
means of pseudopodia, they digest and distribute food, transport oxygen, and
dispose of wastes. Amoebocytes also manufacture the fibers that make up a
sponge's skeleton. In some sponges, these fibers are sharp and spur-like. Other
sponges have softer, more flexible skeletons; we use these pliant, honey-combed
skeletons as bathroom sponges.
Cnidarians
Cnidarians
(phylum Cnidaria) are characterized by radial symmetry and tentacles with
stinging cells. Jellies, sea anemones, hydras, and coral animals are all
cnidarians. Most of the 10,000 cnidarian species are marine.
The basic body
plan of a cnidarian is a sac with a central digestive compartment, the
gastrovascular cavity. A single opening to this cavity functions as both mouth
and anus. This basic body plan has two variations: the sessile polyp and the
floating medusa (Figure 17.11 ). Polyps adhere to the substratum and extend
their tentacles, waiting for prey. Examples of the polyp form are hydras, sea
anemones, and coral animals (Figure 17.12) .A medusa is a flattened, mouth-down
version of the polyp. It moves freely in the water by a combination of passive
drifting and contractions of its bell-shaped body. The animals we generally
call jellies are medusas (jellyfish is another common name, though these
animals are not really fishes, which are vertebrates). Some cnidarians exist
only as polyps, others only as medusas, and still others pass sequentially
through both a medusa stage and a polyp stage in their life cycle.



Figure 17.11 Polyp and medusa forms of cnidarians. Note that
cnidarians have two tissue layers, distinguished in the diagrams by blue and
yellow. The gastrovascular cavity is a digestive sac, meaning that it has only
one opening, which functions as both mouth and anus. {a) Sea anemones are
examples of the polyp form of the basic cnidarian body plan. {b) Jellies are
examples of the medusa form.

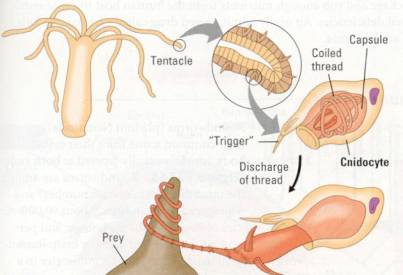
Figure 17.12 Coral animals. Each polyp in this colony is about 3
mm in diameter. Coral animals secrete hard external skeletons of calcium
carbonate (limestone). Each polyp builds on the skeletal remains of earlier
generations to construct the "rocks" we call coral. Though individual
coral animals are small, their collective construction accounts for such
biological wonders as Australia's Great Barrier Reef, which Apollo astronauts
were able to identify from the moon. Tropical coral reefs are home to an
enormous variety of invertebrates and fishes.
Figure 17.13 Cnidocyte action. Each cnidocyte contains a fine
thread coiled within a capsule. When a trigger is stimulated by touch, the
thread shoots out. Some cnidocyte threads entangle prey, while others puncture
the prey and inject a poison.
Cnidarians are
carnivores that use tentacles arranged in a ring around the mouth to capture
prey and push the food into the gastrovascular cavity, where digestion begins.
The undigested remains are eliminated through the mouth/anus. The tentacles are
armed with batteries of cnidocytes ("stinging cells"), unique
structures that function in defense and in the capture of prey (Figure 17.13).
The phylum Cnidaria is named for these stinging cells.
Flatworms
Flatworms (phylum
Platyhelminthes) are [ the simplest bilateral animals. True to their name,
these worms are leaf-like or ribbon-like, ranging from about 1 mm to about 20 m
in length. There are about 20,000 species of flatworms living in marine, fresh
water, and damp terrestrial habitats. Planarians are examples of free-living
(nonparasitic) flatworms (Figure 17.14) .The phylum also includes many
parasitic species, such as flukes and tapeworms.
Parasitic
flatworms called blood flukes are a huge health problem in the tropics. These
worms have suckers that attach to the inside of the blood vessels near the
human host's intestines. This causes a long-lasting disease with such symptoms
as severe abdominal pain, anemia, and dysentery. About 250 million people in 70
countries suffer from blood fluke disease. Flukes generally have complex life
cycles that require more than one host species. People are most commonly
exposed to blood flukes while working in irrigated fields contaminated with
human feces. Blood flukes living in a human host reproduce sexually, and
fertilized eggs pass out in the host's feces. If an egg lands in a pond or
stream, a motile larva hatches. This larva can enter a snail, the next host.
Asexual reproduction in the snail eventually produces other larvae that can
infect humans. A person becomes infected when these larvae penetrate the skin.
Tapeworms
parasitize many vertebrates, including humans. In contrast to planarians and
flukes, most tapeworms have a very long, ribbon like body with repeated parts
(Figure 17.15) .They also differ from other flatworms in not having any
digestive tract at all. Living in partially digested food in the intestines of
their hosts, tapeworms simply absorb nutrients across their body surface. Like
parasitic flukes, tapeworms have a complex life cycle, usually involving more
than one host. Humans can become
infected with tapeworms by eating rare beef containing the worm's larvae. The
larvae are microscopic, but the adults can reach lengths of 20 m in the human
intestine. Such large tapeworms can cause intestinal blockage and rob enough
nutrients from the human host to cause nutritional deficiencies. An orally
administered drug called niclosamide kills the adult worms.
Roundworms
Roundworms
(phylum Nematoda) get their common name from their cylindrical body, which is
usually tapered at both ends (Figure 17.16a).Roundworms are among the most
diverse (in species number) and widespread of all animals. About 90,000 species
of roundworms are known, and perhaps ten times that number actually exist.
Round worms range in length from about a millimeter to a meter. They are found in
most aquatic habitats, in wet soil, and as parasites in the body fluids and
tissues of plants and animals. Free living roundworms in the soil are important
decomposers. Other species are major agricultural pests that attack the roots
of plants, Humans host at least 50 parasitic roundworm species, including
pinworms, hookworms, and the parasite that causes trichinosis (Figure 17.16b).

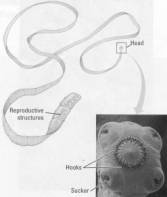
Figure 17.14 Anatomy of a planarian. This free-living flatworm has
a head with two light-detecting eye-spots and a flap at each side that detects
certain chemicals in the water. Dense clusters of nervous tissue form a simple
brain. The digestive tract is highly branched, providing an extensive surface
area for the absorption of nutrients. When the animal feeds, a muscular tube
projects through the mouth and sucks food in. The digestive tract, like that of
cnidarians, is a gastrovascular cavity (a single opening functions as both
mouth and anus). Planarians live on the undersurfaces of rocks in freshwater
ponds and streams.
Figure 17.15 Anatomy of a tapeworm. Humans acquire larvae of these
parasites by eating undercooked meat that is infected. The head of a tapeworm
is armed with suckers and menacing hooks that lock the worm to the intestinal
lining of the host. Behind the head is a long ribbon of units that are little
more than sacs of sex organs. At the back of the worm, mature units containing
thousands of eggs break off and leave the host's body with the feces.
Roundworms
exhibit two evolutionary innovations not found in flat-worms. First, roundworms
have a complete digestive tract, which is a digestive tube with two openings, a
mouth and an anus. This anatomy contrasts with the digestive sac, or
gastrovascular cavity, of cnidarians and flatworms, which uses a single opening
as both mouth and anus. All the remaining animals in our survey of animal phyla
have a complete digestive tract. A complete digestive tract can process food
and absorb nutrients as a meal moves in one direction from one specialized
digestive organ to the next. In humans, for example, the mouth, stomach, and
intestines are examples of digestive organs. A second evolutionary innovation
we see for the first time in roundworms is a body cavity, which in this case is
a pseudo coelom (see Figure 17.8).
Mollusks
Snails and slugs,
oysters and clams, and octopuses and squids are all mollusks (phylum Mollusca).
Mollusks are soft-bodied animals, but most are protected by a hard shell. Slugs, squids, and octopuses have reduced shells,
most of which are internal, or they have lost their she»s completely during
their evolution. Many mollusks feed by using a strap-like rasping organ called
a radula to scrape up food. Garden snails use their radulas like tiny saws to
cut pieces out of leaves. Most of the 150,000 known species of mollusks are
marine, though some inhabit fresh water, and there are land-dwelling mollusks
in the form of snails and slugs.
Despite their
apparent differences, all mollusks have a similar body plan (Figure 17.17) .The
body has three main parts: a muscular foot, usually used for movement; a
visceral mass containing most of the internal organs; and a fold of tissue
called the mantle. The mantle drapes over the visceral mass and secretes the
shell (if one is present).
The three major
classes of mollusks are gastropods (including snails and slugs), bivalves
(including clams and oysters), and cephalopods (including squids and
octopuses). Most gastropods are protected by a single, spiraled shell into
which the animal can retreat when threatened (Figure 17.18a). Many gastropods
have a distinct head with eyes at the tips of tentacles (think of a garden
snail). Bivalves, including numerous species of clams, oysters, mussels, and
scallops, have shells divided into two halves hinged together (Figure 17.18b).
Most bivalves are sedentary, living in sand or mud in marine and freshwater
environments. They use their muscular foot for digging and anchoring.
Cephalopods generally differ from gastropods and sedentary bivalves in being
built for speed and agility. A few cephalopods have large, heavy shells, but in
most the shell is small and internal (as in squids) or missing altogether (as
in octopuses). Cephalopods are marine predators that use beaklike jaws and a
radula to crush or rip prey apart. The cephalopod mouth is at the base of the
foot, which is drawn out into several long tentacles for catching and holding
prey (Figure 17.18c).

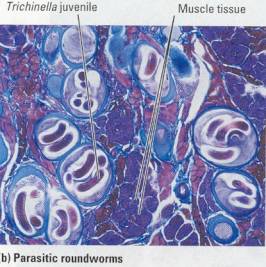
Figure 17.16 Roundworms. (a) This species has the classic
roundworm shape: cylindrical with tapered ends. You can see the mouth at the
end that is more blunt. Not visible is the anus at the other end of a complete
digestive tract. This worm looks like it's wearing a corduroy coat, but the
ridges actually indicate muscles that run the length of the body. (b) The
disease called trichinosis is caused by these roundworms, encysted here in
human muscle tissue. Humans acquire the parasite by eating undercooked pork or
other meat that is infected. The worms then burrow into the human intestine and
eventually travel to other parts of the body, encysting in muscles and other
organs.
Annelids
Annelids (phylum
Annelida) are worms with body segmentation, which is the division of the body
along its length into a series of repeated segments that look like a set of
fused rings. Look closely at an earthworm, an annelid you have all encountered,
and you'll see why these creatures are also called segmented worms (Figure
17.19). In all, there are about 15,000
annelid species, ranging in length from less than 1 mm to the 3-m giant
Australian earthworm. Annelids live in the sea, most freshwater habitats, and
damp soil. The three main classes of annelids are the earthworms and their
relatives, the polychaetes, and the leeches.

Figure 17.17 The general body plan of a mollusk. Note the body
cavity (a true coelom, though a small one) and the complete digestive tract,
with both mouth and anus.
Earthworms eat
their way through the soil, extracting nutrients as the soil passes through the
digestive tube (Figure 17.208). Undigested material, mixed with mucus secreted
into the digestive tract, is eliminated as castings through the anus. Farmers
value earthworms because the animals till the earth, and the castings improve
the texture of the soil. Charles Darwin estimated that each acre of British
farmland had about 50,000 earthworms that produced 18 tons of castings per
year.
In contrast to
earthworms, most polychaetes are marine, mainly crawling or burrowing in the
seafloor. Segmental appendages with hard bristles help the worm wriggle about
in search of small invertebrates to eat. The appendages als-o increase the
animal's surface area for taking up oxygen and disposing of metabolic wastes,
including carbon dioxide (Figure 17.20b ).
The third group
of annelids, leeches, are notorious for the bloodsucking habits of some
species. However, most species are free-living carnivores that eat small
invertebrates such as snails and ins~cts. The majority of leeches live in fresh
water, but a few terrestrial species inhabit moist vegetation in the tropics.
Until the twentieth century, bloodsucking leeches were frequently used by
physicians for bloodletting, the removal of what was considered "bad
blood" from sick patients. Some leeches have razorlike jaws that cut
through the skin, and they secrete saliva containing a strong anesthetic and an
anticoagulant into the wound. The anesthetic makes the bite virtually painless,
and the anticoagulant keeps the blood from clotting. Leech anticoagulant is now
being produced commercially by genetic engineering and may find wide use in
human medicine. Tests show that it prevents blood clots that can cause heart
attacks. Leeches are also still occasionally used to remove blood from bruised
tissues and to help relieve swelling in fingers or toes that have been sewn
back on after accidents (Figure 17.20c). Blood tends to accumulate and cause
swelling in a reattached finger or toe until small veins have a chance to grow
back int9 it. Leeches are applied to remove the excess blood.
Arthropods
Arthropods
(phylum Arthropoda) are named for their Jointed appendages. {such as crabs and
lobsters), arachnids (such as spiders and scorpions), and insects are all
examples of arthropods. Zoologists estimate that the total arthropod population
of Earth numbers about a billion billion (1018) individuals.
Researchers have identified about a million arthropod species, mostly insects.
In fact, two out of every three species of life that have been described are
arthropods. And arthropods are
represented in nearlyall habitats of the biosphere. On the criteria of species
diversity, distribution, and sheer numbers, arthropods must be regarded as the
most successful of all animal phyla.



Figure 17.18 Mollusks. (a) Shell collectors are delighted by the
variety of gastropods. (b) This scallop, a bivalve, has many eyes peering out
between the two halves of the hinged shell. (c) An octopus is a cephalopod
without a shell. All cephalopods have large brains and sophisticated sense
organs, which contribute to the success of these animals as mobile predators. This
octopus lives on the seafloor, where it scurries about in search of crabs and
other food. Its brain is larger and more complex, proportionate to body size,
than that of any other invertebrate. Octopuses are very intelligent and have
shown remarkable learning abilities in laboratory experiments.
General
Characteristics of Arthropods
Arthopods are segmented animals. In contrast to the matching
segments of annelids, however, arthropod segments and their appendages have
become specialized for a great variety of functions. This evolutionary
flexibility contributed to the great diversification of arthropods.
Specialization of segments, or fused groups of segments, also provides for an
efficient division of labor among body regions. For example, the appendages of
different segments are variously modified (for walking, feeding, sensory
reception, copulation, and defense (Figure 17.21).

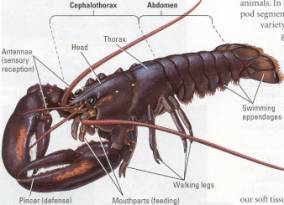
Figure 17.19 Segmented anatomy of an earthworm. Annelids are
segmented both externally and internally. Many of the internal structures are
repeated, segment by segment. The coelom (body cavity) is partitioned by walls
(only two segment walls are fully shown here). The nervous system (yellow)
includes a nerve cord with a cluster of nerve cells in each segment. Excretory
organs (green), which dispose of fluid wastes, are also repeated in each
segment. The digestive tract, however, is not segmented; it passes through the
segment walls from the mouth to the anus. Segmental blood vessels connect
continuous vessels that run along the top (dorsallocation) and bottom (ventral
location) of the worm. The segmental vessels include five pairs of accessory
hearts. The main heart is simply an enlarged region of the dorsal blood vessel
near the head end of the worm.
Figure 17.21 Arthropod characteristics of a lobster. The whole
body, including the appendages, is covered by an exoskeleton. The two distinct
regions of the body are the cephalothorax (consisting of the head and thorax)
and the abdomen. The head bears a pair of eyes, each situated on a movable
stalk. The body is segmented, but this characteristic is only obvious in the
abdomen. The animal has a tool kit of specialized appendages, including
pincers, walking legs, swimming appendages, and two pairs of sensory antennae.
Even the multiple mouthparts are modified legs, which is why they work form
side to side rather than up and down (as our jaws do).
The body of an
arthropod is completely covered by an exoskeleton (external skeleton). This
coat is constructed from layers of protein and a polysaccharide called chitin.
The exoskeleton can be a thick, hard armor over some parts, of the body yet
paper-thin and flexible in other locations, such as the joints. The exoskeleton
protects the animal and provides points of attachment for the muscles that move
the appendages. There are, of course, advantages to wearing hard parts on the
outside. Our own skeleton is interior to most of our soft tissues, an
arrangement that doesn't provide much protection from injury. But our skeleton
does offer the advantage of being able to grow along with the rest of our body.
In contrast, a growing arthropod must occasionally shed its old exoskeleton and
secrete a larger one. This process, called molting, leaves the animal
temporarily vulnerable to predators and other dangers.



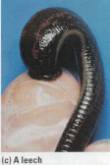
Figure 17.20 Annelids. (a) Giant Australian earthworms are bigger
than most snakes. Perhaps you've slipped on slimy worms, but imagine actually
tripping over one. (b) Polychaetes have segmental appendages that function in
movement and as gills. On the left is a sandworm. The beautiful polychaete on
the right is an example of a fan worm, which lives in a tube it constructs by
mixing mucus with bits of sand and broken shells. Fan worms use their feathery
head-dresses as gills and to extract food particles from the seawater. This
species is called a Christmas tree worm. (c) A nurse applied this medicinal
leech (Hirudo medicinalisl to a patient's sore thumb to drain blood from a
hematoma (abnormal accumulation of blood around an internal injury).
Arthropod
Diversity The four main groups of arthropods are the arachnids, the
crustaceans, the millipedes and centipedes, and the insects. Most arachnids
live on land. Scorpions, spiders, ticks, and mites are examples (Figure 17.22).
Arachnids usually have four pairs of walking legs and a specialized pair of
feeding appendages. In spiders, these feeding appendages are fanglike and
equipped with poison glands. As a spider uses these appendages to immobilize
and chew its prey, it spills digestive juices onto the torn tissues and sucks
up its liquid meal.
Crustaceans are
nearly all aquatic. Crabs, lobsters, crayfish, shrimps, and barnacles are all
crustaceans (Figure 17.23). They all exhibit the crustacean hallmark of
multiple pairs of specialized appendages (see Figure 17.21). One group of
crustaceans, the isopods, is represented on land by pill bugs, which you have
probably found on the undersides of moist leaves and other organic debris.

Figure 17.22 Arachnids. (a) Scorpions are nocturnal hunters. Their
ancestors were among the first terrestrial carnivores, preying on herbivorous
arthropods that fed on the early land plants. Scorpions have a pair of
appendages modified as large pincers that function in defense and food capture.
The tip of the tail bears a poisonous stinger. Scorpions eat mainly insects and
spiders. They will sting people only when prodded or stepped on. (If you camp
in the desert and leave your shoes on the ground when you go to bed, make sure
there are no scorpions in those shoes before putting them on in the morning.)
(b) Spiders are usually most active during the daytime, hunting insects or
trapping them in webs. Spiders spin their webs of liquid silk, which solidifies
as it comes out of specialized glands. Each spider engineers a style of web
that is characteristic of its species, getting the web right on the very first
try. Besides building their webs of silk, spiders use the fibers in many other
ways: as droplines for rapid escape; as cloth that covers eggs; and even as
"gift wrapping" for food that certain male spiders offer to seduce
females. (c) This magnified house dust mite is a ubiquitous scavenger in our
homes. Each square inch of carpet and every one of those dust balls under a bed
s are like cities to thousands of dust mites. Unlike some mites that carry
pathogenic bacteria, dust mites are harmless except to people who are allergic
to the mites' feces.



(a) A shrimp
(b) Barnacles Figure
17.24 A millipede
Figure 17.23 Crustaceans. (a) A grass shrimp. (b) Easily confused
with bivalve mollusks, barnacles are actually sessile crustaceans with
exoskeletons hardened into shells by calcium carbonate (lime). The jointed
appendages projecting from the shell capture small plankton.
Millipedes and
centipedes have similar segments over most of the body and superficially
resemble annelids, but their jointed legs give them away as arthropods.
Millipedes are landlubbers that eat decaying plant matter (Figure 17.24) .They
have two pairs of short legs per body segment. Centipedes are terrestrial
carnivores, with a pair of poison claws used in defense and to paralyze prey,
such as cockroaches and flies. Each of their body segments bears a single pair
of long legs.
In species
diversity, insects outnumber all other forms of life combined. They live
in almost every terrestrial habitat and in fresh water, and flying insects fill
the air. Insects are rare, though not absent, in the seas, where crustaceans
are the dominant arthropods. There is a whole big branch of biology, called
entomology. that specializes in the study of insects.
The oldest insect
fossils date back to about 400 million years ago, during the Paleozoic era (see
Table 14.1). Later, the evolution of flight sparked an explosion in insect
variety (Figure 17.25). Like the grasshopper in Figure 17.25, most insects have
a three-part body: head, thorax, and abdomen. The head usually bears a pair of
sensory antennae and a pair of eyes. Several pairs of mouthparts are adapted
for particular kinds of eating-for example, for biting and chewing-plant
material in grasshoppers; for lapping up fluids in houseflies; and for piercing
skin and sucking blood in ! mosquitoes and other biting flies. Most adult
insects have three pairs of legs and one or two pairs of wings, all borne on
the thorax.
Flight is
obviously one key to the great success of insects. An animal that can fly can
escape many predators, find food and mates, and disperse to new habitats much
faster than an animal that must crawl about on the ground. Because their wings
are extensions of the exoskeleton and not true appendages, insects can fly
without sacrificing any walking legs. By contrast, the flying vertebrates-birds
and bats-have one of their two pairs of wa1king legs modified for wings, which
explains why these vertebrates are generally not very swift on the ground.
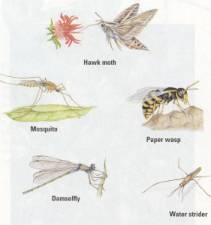
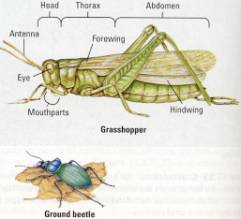

Figure 17.25 A small
sample of insect diversity
Figure 17.26 Metamorphosis
of a monarch butterfly. (a) The larva (caterpillar) spends its time eating and
growing, molting as it grows. (b) After several molts, the larva encases itself
in a cocoon and becomes a pupa. (c) Within the pupa, the larval organs break
down and adult organs develop from cells that were dormant in the larva. (d)
Finally, the adult emerges from the cocoon. (e) The butterfly flies off and
reproduces, nourished mainly from the calories it stored when it was a
caterpillar.
Many insects
undergo metamorphosis in their development. In the case of grasshoppers and some
other insect groups, the young resemble adults but are smaller and have
different body proportions. The animal goes through a series of molts, each
time looking more like an adult, until it reaches full size. In other cases,
insects have distinctive larval stages specialized for eating and growing that
are known by such names as maggots, grubs, or caterpillars. The larval stage
looks entirely different from the adult stage, which is specialized for
dispersal and reproduction. Metamorphosis from the larva to the adult occurs
during a pupal stage (Figure 17.26).
Animals so
numerous, diverse, and widespread as insects are bound to affect the lives of
all other terrestrial organisms, including humans. On one hand, we depend on
bees, flies, and many other insects to pollinate our crops and orchards. On the
other hand, insects are carriers of the micro-organisms that cause many
diseases, including malaria and African sleeping sickness. Insects also compete
with humans for food. In parts of Africa, for instance, insects claim about 75%
of the crops. Trying to minimize their losses, farmers in the United States
spend billions of dollars each year on pesticides, spraying crops with massive
doses of some of the deadliest poisons ever invented. Try as they may, not even
humans have challenged the preeminence of insects and their arthropod kin. As
Cornell University's Thomas Eisner puts it: "Bugs are not going to inherit
the Earth. They own it now. So we might as well make peace with the landlord:
Echinoderms
The echinoderms
.(phylum Echinodermata) are named for their spiny surfaces. Sea urchins, the
porcupines of the invertebrates, are certainly echinoderms that live up to the
phylum name. Among the other echmoderms are sea stars, sand dollars, and sea
cucumbers (Figure 17.27). Echinoderms
are all marine. Most are sessile or slow moving. Echinoderms lack body
segments, and most have radial symmetry as adults. Both the external and the
internal parts of a sea star, for instance, radiate from the center like spokes
of a wheel. In contrast to the adult, the larval stage of echinoderms is
bilaterally symmetrical. This supports other evidence that echinoderms are not
closely related to cnidarians or other radial animals that never show bilateral
symmetry. Most echinoderms have an endoskeleton (interior skeleton) constructed
from hard plates just beneath the skin. Bumps and spines of this endoskeleton
account for the animal's rough or prickly surface. Unique to echinoderms is the
water vascular system, a network of water-filled canals that circulate water
throughout the echinoderm's body, facilitating gas exchange and waste disposal.
The water vascular system also branches into extensions called tube feet. A sea
star or sea urchin pulls itself slowly over the seafloor using its suction-
cup-like tube feet. Sea stars also use their tube feet to grip prey during
feeding (see Figure 17.27a).
Looking at sea
stars and other adult echinoderms, you may think they .have little in common
with humans and other vertebrates. But if you return to Figure 17.6, you'll see
that echinoderms share an evolutionary branch with chordates, the phylum that
includes vertebrates. Analysis of embryonic development reveals this
relationship. The mechanism of coelom formation and many other details of embryology
differentiate the echinoderms and chordates from the evolutionary branch that
includes mollusks, annelids, and arthropods. With this phylogenetic context,
we're now ready to make the transition from invertebrates to vertebrates.

Figure 17.27 Echinoderms. (a) The mouth of a sea star, not visible
here, is located in the center of the undersurface. The inset shows how the
tube feet function in feeding. When a sea star encounters an oyster or clam,
its favorite foods, it grips the mollusk's shell with its tube feet and
positions its mouth next to the narrow opening between the two halves of the
prey's shell. The sea star then pushes its stomach out through its mouth and
through the crack in the mollusk's shell. The predator then digests the soft
tissue of its prey. (b) In contrast to sea stars, sea urchins are spherical and
have no arms. If you look closely, you can see the long tube feet projecting
among the spines. Unlike sea stars, which are mostly carnivorous, sea urchins
mainly graze on seaweed and other algae. (c) On casual inspection, sea
cucumbers do not look much like other echinoderms. Sea cucumbers lack spines,
and the hard endoskeleton is much reduced. However, closer inspection reveals
many echinoderm traits, including five rows of tube feet.
The
vertebrate Genealogy
Most of us are curious about our
genealogy. On the personal level, we wonder about our family ancestry. As
biology students, we are interested in tracing human ancestry within the
broader scope of the entire animal kingdom. In this quest, we ask three
questions: What were our ancestors like? How are we related to other animals?
and What are our closest relatives?



Figure 17.28 Backbone, extra long. Vertebrates are named for their
backbone, which consists of a series of vertebrae. The vertebrate hallmark is
apparent in this snake skeleton. You can also see the skull, the bony case
protecting the brain. The backbone and skull are parts of an endoskeleton, a
skeleton inside the animal rather than covering it.
Figure 17.29 Invertebrate chordates. (a) lancelets owe their name
to their bladelike shape. Marine animals only a few centimeters long, lancelets
wiggle backward into sand, leaving only their head exposed. The animal filters
tiny food particles from the seawater. (b) This adult tunicate, or sea squirt,
is a sessile filter feeder that bears little resemblance to other chordates.
However, a tunicate goes through a larval stage that is unmistakably chordate.
Figure 17.30 Cordate
characteristics
In this section, we trace the evolution of
the vertebrates, the group that includes humans and their closest relatives.
Mammals, birds, reptiles, amphibians, and the various classes of fishes are all
classified as vertebrates. Among the unique vertebrate features are the cranium
and backbone, a series of vertebrae for which the group is named (Figure
17.28). Our first step in tracing the vertebrate genealogy is to determine
where vertebrates fit in the animal kingdom.
Characteristics
of Chordates
Vertebrates make
up one subphylum of the phylum Chordata. Our phylum also includes two subphyla
of invertebrates, animals lacking a backbone: lancelets and tunicates (Figure
17.29). These invertebrate chordates and vertebrates all share four key
features that appear in the embryo and sometimes in the adult. These four
chordate hallmarks are ( 1) a dorsal, hollow nerve cord ( the chordate brain
and spinal cord); (2) a notochord, which is a flexible, longitudinal rod
located between the digestive tract and the nerve cord; ( 3) pharyngeal slits,
which are gill structures in the pharynx, the region of the digestive tube just
behind the mouth; and ( 4) a post -anal tail, which is a tail to the rear of
the anus (Figure 17.30). Though these chordate characteristics are often
difficult to recognize in the adult animal, they are always present in chordate
embryos. For example, the notochord, for which our phylum is named, persists in
adult humans only in the form of the cartilage disks that function as cushions
between the vertebrae. (Back injuries described as "ruptured disks"
or "slipped disks" refer to these structures. )
Body segmentation
is another chordate characteristic, though not a unique one. The chordate
version of segmentation probably evolved independently of the segmentation we
observe in annelids and arthropods. Chordate segmentation is apparent in the
backbone of vertebrates (see Figure 17.28) and in the segmental muscles of all
chordates (see the chevron-shaped- »» -muscles in the lancelet of Figure
17.29a). Segmental musculature is not so obvious in adult humans unless one is
motivated enough to sculpture those washboard "abs of steel."
Vertebrates
retain the basic chordate characteristics, but have additional features that
are unique, including, of course, the backbone (see Figure 17.28). Figure 17.31
is an overview of chordate and vertebrate evolution that will provide a context
for our survey of the vertebrate classes.
Fishes
The first
vertebrates probably evolved during the early Cambrian period about 540 million
years ago. These early vertebrates, the agnathans, lacked jaws. Agnathans are
represented today by vertebrates called lampreys (see Figure 17.31). Some
lampreys are parasites that use their jawless mouths as suckers to attach to
the sides of large fishes and draw blood. In contrast, most vertebrates have
jaws, which are hinged skeletons that work the mouth. We know from the fossil
record that the first jawed vertebrates

were fishes that replaced most agnathans by about 400 million
years ago. In addition to jaws, these fishes had two pairs of fins, which made
them maneuverable swimmers. Some of those fishes were more than 10 m long.
Sporting jaws and fins, some of the early fishes were active predators that
could chase prey and bite off chunks of flesh. Even today, most fishes are
carnivores. The two major groups of living fishes are the class Chondrichthyes
( cartilaginous fishes-the sharks and rays) and the class Osteichthyes (the
bony fishes, including such familiar groups as tuna, trout, and goldfish).
Cartilaginous
fishes have a flexible skeleton made of cartilage. Most sharks are adept
predators-fast swimmers with streamlined bodies, acute senses, and powerful
jaws (Figure 17.]2a). A shark does not have keen eyesight, but its sense of
smell is very sharp. In addition, special electrosensors on the head can detect
minute electrical fields produced by muscle contractions in nearby animals.
Sharks also have a lateral line system. a row of sensory organs running along
each side of the body. Sensitive to changes in water pressure, the lateral line
system enables a shark to detect minor vibrations caused by animals swimming in
its neighborhood. There are fewer than 1,000 living species of cartilaginous
fishes, nearly all of them marine.
Bony fishes
(Figure 17.]2b) have a skeleton reinforced by hard calcium salts. They also
have a lateral line system, a keen sense of smell, and excellent eyesight. On
each side of the head, a protective flap called the operculwn (plural,
opercula) covers a chamber housing the gills. Movement of the operculum allows
the fish to breathe without swimming. (By contrast, sharks lack opercula and
must swim to pass water over their gills. ) Bony fishes also have a specialized
organ that helps keep them buoyant-the swim bladder. a gasfilled sac. Thus,
many bony fishes can conserve energy by remaining almost motionless, in
contrast to sharks, which sink to the bottom if they stop swimming. Some bony
fishes have a connection between the swim bladder and the digestive tract that
enables them to gulp air and extract oxygen from it when the dissolved oxygen
level in the water gets too low. In fact, swim bladders evolved from simple
lungs that augmented gills in absorbing oxygen from the water of stagnant
swamps, where the first bony fishes lived.


Figure 17.32 Two classes of fishes. (a) A member of the class
Chondrichthyes, the cartilaginous (b) A member of the class Osteichthyes, the
bony fishes.
The largest class
of vertebrates { about 30,000 species ), bony fishes are common in the seas and
in freshwater habitats. Most bony fishes, including trout, bass, perch, and
tuna, are ray-finned fishes. Their fins are supported by thin, flexible
skeletal rays (see Figure 17 .32b). A second evolutionary branch of bony fishes
includes the lungfishes and lobe-finned fishes. Lung-fishes live today in the
Southern Hemisphere. They inhabit stagnant ponds and swamps, surfacing to gulp
air into their lungs. The lobe-fins are named for their muscular fins supported
by stout bones. Lobe-fins are extinct except for one species, a deep-sea
dweller that may use its fins to waddle along the seafloor. Ancient freshwater
lobe-finned fishes with lungs played a key role in the evolution of amphibians,
the first terrestrial vertebrates.
Amphibians
In Greek, the,
word amphibios means "living a double life. Most members of the class
Amphibia exhibit a mixture of aquatic and terrestrial adaptations. Most species
are tied to water because their eggs, lacking shells, dry out quickly in the
air; The frog in Figure 17.33 spends much of its time on land, but it lays its
eggs in water. An egg develops into a larva called a tadpole, a legless,
aquatic algae-eater with gills, a lateral line system resembling that of
fishes, and a long finned tail. In changing into a frog, the tadpole undergoes
a radical metamorphosis. When a young frog crawls onto shore and begins life as
a terrestrial insect -eater, it has four legs, air-breathing lungs instead of
gills, a pair of external eardrums, and no lateral line system. Because of
metamorphosis, many amphibians truly live a double life. But even as adults,
amphibians are most abundant in damp habitats, such as swamps and rain forests.
This is partly because amphibians depend on their moist skin to supplement lung
function in exchanging gases with the environment. Thus, even those frogs that
are adapted to relatively dry habitats spend much of their time in humid
burrows or under piles of moist leaves. The amphibians of today, including
frogs and salamanders, account for only about 8% of all living vertebrates, or about
4,000 species.


Figure 17.33 The uduallifeu of an amphibian. Though not all
amphibians have aquatic larval stages. this class of vertebrates is named for
the familiar tadpole-to-frog metamorphosis of many species

Figure 17.34 The origin of tetrapods. Fossils of some lobe-finned
fishes have skeletal supports extending into their fins Early amphibians left
fossilized limb skeletons that probably functioned in movement on land
Amphibians were
the first vertebrates to colonize land. They descended from fishes that liad
lungs and fins with muscles and skeletal supports strong enough to enable some
movement, however clumsy, on land (Figure 17.34). The fossil record chronicles
the evolution of four-limbed amphibians from fishlike ancestors. Terrestrial vertebrates-amphibians,
reptiles, birds, and mammals-are collectively called tetrapods, which means
"four legs:' Had our amphibian ancestors had three pairs of legs on their
undersides instead of just two, we might be hexapods. This image seems silly,
but serves to reinforce the point that evolution, as descent with modification,
is constrained by history.
Reptiles
Class Reptilia
includes snakes, lizards, turtles, crocodiles, and alligators. The evolution of
reptiles from an amphibian ancestor paralleled many additional adaptations for
living on land. Scales containing a protein called keratin waterproof the skin
of a reptile, helping to prevent dehydration in dry air. Reptiles cannot
breathe through their dry skin and obtain most of their oxygen with their
lungs. Another breakthrough for living on land that evolved in reptiles is the
amniotic egg, a water-containing egg enclosed in a shell (Figure 17.35). The
amniotic egg functions as a "self -contained pond" that enables
reptiles to complete their life cycle on land. With adaptations such as
waterproof skin and amniotic eggs, reptiles broke their ancestral ties to
aquatic habitats. There are about 6,500 species of reptiles alive today.
Reptiles are
sometimes labeled "cold-blooded" animals because they do not use
their metabolism extensively to control body temperature. But reptiles do
regulate body temperature through behavioral adaptations. For example, many
lizards regulate their internal temperature by basking in the sun when the air
is cool and seeking shade when the air is too warm. Because they absorb
external heat rather than generating much of their own, reptiles are said to be
ectotherms, a term more accurate than cold-blooded. By heating directly with
solar energy rather than through the metabolic breakdown of food, a reptile can
survive on less than 10% of the calories required by a mammal of equivalent
size.
As successful as
reptiles are today, they were far more widespread, numerous, and diverse during
the Mesozoic era, which ,is sometimes called the "age of reptiles:'
Reptiles diversified extensively during that era, producing a dynasty that
lasted until the end of the Mesozoic, about 65 million years ago. Dinosaurs,
the most diverse group, included the largest animals ever to inhabit land. Some
were gentle giants that lumbered about while browsing vegetation. Others were
voracious carnivores that chased their larger prey on two legs (Figure 17.36).


Figure 17.35 Terrestrial equipment of reptiles. This bull snake
displays two reptilian adaptations to living on land: a waterproof skin with
keratinized scales; and amniotic eggs, with shells that protect a watery,
nutritious internal environment where the embryo can develop on land. Snakes
evolved from lizards that adapted to a burrowing lifestyle.
Figure 17.36 A Mesozoic feeding frenzy. Hunting in packs, Deinonychus (meaning "terrible claw")
probably used its sickle-shaped claws to slash at larger prey.


Figure 17.37 A bald eagle in flight. Bird wings are airfoils,
which have shapes that create lift by altering air currents. Air passing over a
wing must travel farther in the same amount of time than air passing under the
wing. This expands the air above the wing relative to the-air below the wing.
And this makes the air pressure pushing upward against the lower wing surface
greater than the pressure of the expanded air pushing downward on the wing. The
wings of birds and airplanes owe their "lift" to this pressure
differential.
The age of reptiles began to wane about 70
million years ago. During the Cretaceous, the last period of the Mesozoic era,
global climate became cooler and more variable. This was a period of mass
extinctions that claimed all the dinosaurs by about 65 million years ago,
except for one lineage. That lone surviving lineage is represented today by
birds.
Birds
Birds (class
Aves)evolved during the great reptilian radiation of the Mesozoic era. Amniotic eggs and scales on the legs
are just two of the reptilian features
we see in birds. But modern birds look quite different from modern reptiles
because of their feathers and other distinctive flight equipment. Almost all of
the 8,600 living bird species are airborne. The few flightless species,
including the ostrich, evolved from flying ancestors. Appreciating the avian
world is all about understanding flight.
Almost every
element of bird anatomy is modified in some way that enhances flight. The bones
have a honeycombed structure that makes them strong but light ( the wings of
airplanes have the same basic construction).
For example, a huge seagoing species called the frigate bird has a
wingspan of more than 2 m, but its whole skeleton weighs only about 113 g (4
ounces). Another adaptation that reduces the weight of birds is the absence of
some internal organs found in other vertebrates. Female birds, for instance,
have only one ovary instead of a pair. Also, modern birds are toothless, an
adaptation that trims the weight of the head (no uncontrolled nosedives). Birds
do not chew food in the mouth but grind it in the gizzard, a chamber of the
digestive tract near the stomach.
Flying requires a
great expenditure of energy and an active metabolism. In contrast to the
ectothermic reptiles, birds are endotherms. That means they use their own
metabolic heat to maintain a warm, constant body temperature. A bird's most
obvious flight equipment is its wings. Bird wings are air-foils that illustrate
the same principles of aerodynamics as the wings of an airplane (Figure 17.37).
A bird's flight motors are its powerful breast muscles, which are anchored to a
keel-like breastbone. It is mainly these flight muscles that we call
"white meat" on a turkey or chicken. Some birds, such as eagles and
hawks, have wings adapted for soaring on air currents and flap their wings only
occasionally. Other birds, including hummingbirds, excel at maneuvering but
must flap continuously to stay aloft. In either case, it is the shape and
arrangement of the feathers that form the wing into an air-foil. Feathers are
made of keratin, the same protein that forms our hair and fingernails as well
as the scales of reptiles. Feathers may have functioned first as insulation,
helping birds retain body heat, only later being co-opted as flight gear.
In tracing the
ancestry of birds back to the Mesozoic era, we must search for the oldest
fossils with feathered wings that could have the functioned in flight. Fossils
of an ancient bird named Archaeopteryx have been found in Bavarian limestone in
Germany and date back some 150 million years into the dinosaurs, birds, and
Jurassic period (see Figure 14.15). Archaeopteryx is not considered the
ancestor of modern birds, and paleontologists place it on a side branch of the
avian linage Nonetheless, Archaeopteryx probably was derived from ancestral
forms that also gave rise to modern birds. Its skeletal anatomy indicates that
it was a weak flyer, perhaps mainly a tree-dwelling glider. A combination of
gliding downward and jumping into the air from the ground may have been the
earliest mode of flying in the bird lineage.


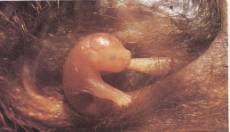
Figure 17.38 The three major groups of mammals. (a) Monotremes,
such as this echidna, are the only mammals that lay eggs (inset). (b) The young
of marsupials, such as this brushtail opossum, are born very early in their
development. They finish their growth while nursing from a nipple in their
mother's pouch. (c) In eutherians (placentals), such as these zebras, young
develop within the uterus of the mother. There they are nurtured by the flow of
blood though the dense network of vessels in the placenta. The reddish portion
of the afterbirth clinging to the newborn zebra in this photograph is the
placenta. (see prior)
Mammals
Mammals (class
Mammalia) evolved from reptiles about 225 million years ago, long before there
were any dinosaurs. During the peak of the age of reptiles, there were a number
of mouse-sized, nocturnal mammals that lived on a diet of insects. Mammals
became much more diverse after the downfall of the dinosaurs. Most mammals are
terrestrial. However, there are nearly 1,000 species of winged mammals, the
bats. And about 80 species of dolphins, porpoises, and whales are totally
aquatic. The blue whale, an endangered species that grows to lengths of nearly
30 m, is the largest animal that has ever existed.
Two features-hair
and mammary glands that produce milk that nourishes the young-are mammalian
hallmarks. The main function of hair is to insulate the body and help maintain
a warm, constant internal temperature; mammals, like birds, are endotherms.
There are three major groups of mammals: the monotremes, the marsupials, and
theeutherians. The duck-billed platypus is one of only threeexisting species of
monotremes, the egg-laying mammals. The platypus lives along rivers in eastern
Australia and on the nearby island of Tasmania. It eats mainly small shrimps
and aquatic insects. The female usually lays two eggs and incubates them in a
leaf nest. After hatching, the young nurse by licking up milk secreted onto the
mother's fur. The animals called echidnas are also monotremes (Figure 17.38a).
Most mammals are
born rather than hatched. During gestation in marsupials and eutherians, the
embryos are nurtured inside the mother by an organ called the placenta.
Consisting of both embryonic and maternal tissues, the placenta joins the
embryo to the mother within the uterus. The embryo is nurtured by maternal
blood that flows close to the embryonic blood system in the placenta. The
embryo is bathed in fluid contained by an amniotic sac, which is homologous to
the fluid compartment within the amniotic eggs of reptiles.
Marsupials are
the so-called pouched mammals, including kangaroos and koalas. These mammals
have a brief gestation and give birth to tiny, embryonic offspring that
complete development while attached to the mother's nipples. The nursing young
are usually housed in an external pouch, called the marsupium, on the mother's
abdomen (Figure 17.38b ). Nearly all marsupials live in Australia, New Zealand,
and Central and South America. Australia has been a marsupial sanctuary for
much of the past 60 million years. Australian marsupials have diversified
extensively, filling terrestrial habitats that on other continents are occupied
by eutherian mammals.
Eutherians are
also called placental mammals because their placentas provide more intimate and
long-lasting association between the mother and her developing young than do
marsupial placentas (Figure 17.38c ) .Eutherians make up almost 95% of the
4,500 species of living mammals. Dogs, cats, cows, rodents, rabbits, bats, and
whales are all examples of eutherian mammals. One of the eutherian groups is
the order Primates, includes monkeys, apes, and humans.

Figure 17.39 The arboreal athleticism of primates. This orangutan
displays primate adaptations for living in the trees: limber shoulder joints,
manual dexterity, and stereo vision due to eyes on the front of the face.
The
Human Ancestry
We have now traced the animal genealogy to
the mammalian group that includes Homo sapiens and its closest kin. We are
primates. To understand .what that means, we must trace our ancestry back to
the trees, where some of our most treasured traits originated as arboreal
adaptations.
The
Evolution of Primates
Primate evolution provides a context for
understanding human origins. The fossil record supports the hypothesis that
primates evolved from insect-eating mammals during the late Cretaceous period,
about 65 million years ago. Those early primates were small, arboreal {
tree-dwelling) mammals. Thus, the order
Primates was first distinguished by characteristics that were shaped, through
natural selection, by the demands of living in the trees. For example, primates have limber shoulder
joints, which make it possible to brachiate {swing from one branch to another).
The dexterous hands of primates can hang on to branches and manipulate food.
Nails have replaced claws in many primate species, and the fingers are very
sensitive. The eyes of primates are close together on the front of the face.
The overlapping fields of vision of the two eyes enhance depth perception, an
obvious advantage when brachiating. Excellent eye-hand coordination is also
important for arboreal maneuvering {Figure 17.39). Parental care is essential
for young animals in the trees. Mammals devote more energy to caring for their
young than most other vertebrates, and primates are among the most attentive
parents of all mammals. Most primates have single births and nurture their
offspring for a long time. Though humans do not live in trees, we retain in
modified form many of the traits that originated there.
Taxonomists divide the primates into twp
main groups: prosimians and anthropoids. The oldest primate fossils are
prosimians. Modern prosimians include the lemurs of Madagascar and the lorises,
pottos, and tarsiers that live in tropical Africa and southern Asia (Figure
17.40a). Anthropoids include monkeys, apes, and humans. All monkeys in the New
World ( the Americas) are arboreal and are distinguished by prehensile tails
that function as an extra appendage for brachiating (Figure 17.40b). (If you
see a monkey in a zoo swinging by its tail, you know it's from the New World. )
Although some Old World monkeys are also arboreal, their tails are not
prehensile. And many Old World monkeys, including baboons, macaques, and
mandrills, are mainly ground-dwellers (Figure 17.40c).








Figure
17.40 Primate diversity (a) A
prosimian. (b) – (c) Monkeys (d) – (g) Apes
(h) human
Our closest
anthropoid relatives are the apes: gibbons, orangutans, gorillas, and
chimpanzees (Figure 17.40d-g). Modern apes live only in tropical regions of the
Old World. With the exception of some gibbons, apes are larger than monkeys,
with relatively long arms and short legs and no tail. Although all the apes are capable of brachiation,
only gibbons and orangutans are primarily arboreal. Gorillas and chimpanzees
are highly social. Apes have larger brains proportionate to body size than
monkeys, an their behavior is consequently more adaptable.
The Emergence of Humankind
Humanity
is one very young twig on the vertebrate branch, just one of many branches on
the tree of life. In the continuum of life spanning 3.5 billion years, humans
and apes have shared a common ancestry for all but the last 5-7 million years
(Figure 17.41 ). Put another way, if we compressed the history of life to a
year, humans and chimpanzees diverged from a common ancestor less than 18 hours
ago. The fossil record and molecular systematics concur in that vintage for the
human lineage. Paleoanthropology, the study of human evolution, focuses on this
very thin slice of biological history. Some Common Misconceptions Certain
misconceptions about human evolution that were generated during the early part
of the twentieth century still persist today in the minds of many, long after
these myths have been debunked by the fossil evidence.

Figure
17.41 Primary Phylogeny
Let's first
dispose of the myth that our ancestors were chimpanzees or any other modern
apes. Chimpanzees and humans represent two divergent branches of the anthropoid
tree that evolved from a common, less specialized ancestor. Chimps are not our
parent species, but more like our phylogenetic siblings or cousins.
Another
misconception envisions human evolution as a ladder with a series of steps
leading directly from an ancestral anthropoid to Homo sapiens. This is often
illustrated as a parade of fossil hominids (members of the human family)
becoming progressively more modern as they march across the page. If human
evolution is a parade, then it is a disorderly one, with many splinter groups
having traveled down dead ends. At times in hominid history, several different
human species coexisted (Figure 17.42). Human phylogeny is more like a
multibranched bush than a ladder, with our species being the tip of the only
twig that still lives.
One more myth we
must bury is the notion that various human characteristics, such as upright
posture and an enlarged brain, evolved in unison. A popular image is of early
humans as half-stooped, half-witted cave-dwellers. In fact, we know from the fossil record that different human features
evolved at different rates, with erect posture, or bipedalism, leading the way.
Our pedigree includes ancestors who walked upright but had ape-sized brains.
After dismissing some of the folklore on human evolution, however, we must
admit that many questions about our ancestry have not yet been
resolved.
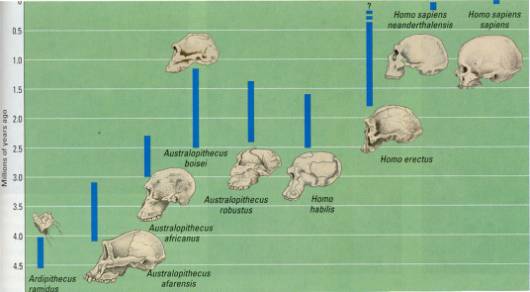
Figure 17.42 A timeline of human evolution. Notice that there have been times when two
or more hominid species coexisted. The skulls are all drawn to the same scale
enable you to compare the sizes of craniums and hence brain
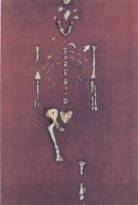

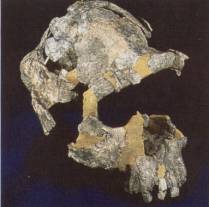
Figure 17.43 The antiquity of upright posture. (a) Lucy, a
3.18-million-year-old skeleton, represents the hominid species Australopithecus
afarensis. Fragments of the pelvis and skull put A. afarensis on two feet. (b)
Some 3.7 million years ago, several bipedal (upright-walking) hominids left
footprints in damp volcanic ash in what is now Tanzania in East Africa. The
prints fossilized and were discovered by British anthropologist Mary Leakey in
1978. The footprints are part of the strong evidence that bipedalism is a vefry
old human trait. (c) This A afarensis
skull, 3.9 million years old, articulated with a vertical back bone. The
upright posture of humans is a least that old.
Australopithecus
and the Antiquity of Bipedalism Before there was Homo, several hominid
species of the genus Australopithecus walked the African savanna (grasslands
with clumps of trees ). Paleoanthropologists have focused much of their
attention on A. afarensis, an early species of Australopithecus. Fossil
evidence now pushes bipedalism in A. afarensis back to at least 4 million years
ago (Figure 17.43) .Doubt remains whether an even older hominid, Ardipithecus
ramidus, which dates back at least 4.4 million years, was bipedal.
One of the most
complete fossil skeletons of A. afarensis dates to about 3.2 million years ago
in East Africa. Nicknamed Lucy by her discoverers, the individual was a female,
only about 3 feet tall, with a head about the size of a softball (see Figure
17.43a). Lucy and her kind lived in savanna areas and may have subsisted on
nuts and seeds, bird eggs, and whatever animals they could catch or scavenge
from kills made by more efficient predators such as large cats and dogs.
All
Australopithecus species were extinct by about 1.4 million years ago. Some of
the later species overlapped in time with early species of our own genus, Homo
( see Figure 17.42 ) .Much debate centers on the evolutionary relationships of
the Australopithecus species to each other and to Homo. Were the Australopithecus
species all evolutionary side branches? Or were some of them ancestors to later
humans? Either way, these early hominids show us that th~ fundamental human
trait ofbipedalism evolved millions of years before the other major human
trait-an enlarged brain. As evolutionary biologist Stephen Jay Gould put it,
"Mankind stood up first and got smart later”.
Homo habilis
and the Evolution of Inventive Minds Enlargement of the human brain is
first evident in fossils from East Africa dating to the latter part of the era
of Australopithecus, about 2.5 million years ago. Anthropologists have
found skulls with brain capacities intermediate in size between those of the
latest Australopithecus species and those of Homo sapiens. Simple handmade
stone tools are sometimes found with the larger-brained fossils, which have
been dubbed Homo habilis ("handy man"). After walking upright for
about 2 million years, humans were finally beginning to use their manual
dexterity and big brains to invent tools that enhanced their hunting,
gathering, and scavenging on the African savanna.
Homo erectus and the
Global Dispersal of Humanity The first species to extend humanity's
range from its birthplace in Africa to other continents was Homo erectus,
perhaps a descendant of H. habilis. The global dispersal began about 1.8
million years ago. But don't picture this migration as a mad dash for new
territory or even as a casual stroll. If H. erectus simply expanded its range
from Africa by about a mile per year, it would take only about 12,000 years to
populate many regions of Asia and Europe. The gradual spread of humanity may
have been associated with a change in diet to include a larger proportion of
meat. In general, animals that hunt require more geographic territory than
animals that feed mainly on vegetation.
Homo erectus
was taller than H. habilis and had a larger brain capacity. During the
1.5 million years the species existed, the H. erectus brain increased to as
large as 1,200 cubic centimeters ( cm3), a brain capacity that overlaps the
normal range for modern humans. Intelligence enabled humans to continue
succeeding in Africa and also to survive in the colder climates of the north.
Homo erectus resided in huts or caves, built fires, made clothes from animal
skins, and designed stone tools that were more refined than the tools of H.
habilis. In anatomical and physiological adaptations, H. erectus was poorly
equipped for life outside the tropics but made up for the deficiencies with
cleverness and social cooperation.
Some African,
Asian, European, and Australasian ( from Indonesia, New Guinea, and Australia)
populations of H. erectus gave rise to regionally diverse descendants that had
even larger brains. Among these descendants of H. erectus were the
Neanderthals, who lived in Europe, the Middle East, and parts of Asia from
about 130,000 years ago to about 35,000 years ago. (They are named Neanderthals
because their fossils were first found in the Neander Valley of Germany. )
Compared with us, Neanderthals had slightly heavier browridges and less
pronounced chins, but their brains, on average, were slightly larger than ours.
Neanderthals were skilled toolmakers, and they participated in burials and
other rituals that required abstract thought. Much current research on the Neanderthal
skull addresses an intriguing question: Did Neanderthals have the anatomical
equipment necessary for speech?
The Origin of Homo
sapiens The oldest known post-H. erectus fossils, dating back over
300,000 years, are found in Africa. Many paleoanthropologists group these
African fossils along with Neanderthals and various Asian and Australasian
fossils as the earliest forms of our species, Homo sapiens. These regionally
diverse descendants of H. erectus are sometimes referred to as "archaic Homo
sapiens." The oldest fully modern fossils of H. sapiens-skulls and human
evolution. other bones that look essentially like those of today's humans-are
about 100,000 years old and are located in Africa. Similar fossils almost as
ancient have also been discovered in caves in Israel. The famous fossils of
modern humans from the Cro-Magnon caves of France date back about 35,000 years.
The relationship
of the Cro-Magnons and other modern humans to archaic Homo sapiens is a
question that will continue to engage paleoanthropologists for decades. What
was the fate of the various descendants of Homo erectus who populated different
parts of the world! In the view of some anthropologists, these archaic H.
sapiens populations gave rise to modern humans. According to this hypothesis,
called the multiregional hypothesis, modern humans evolved simultaneously in
different parts of the world (Figure 17.44a). If this view is correct, then the
geographic diversity of humans originated relatively early, when H. erectus
spread from Africa into the other continents between 1 and 2 million years ago.
This hypothesis accounts for the great genetic similarity of all modern people
by pointing out that interbreeding among neighboring populations has always
provided corridors for gene flow throughout the entire geographic range of
humanity.
In sharp contrast
to the multiregional hypothesis is a hypothesis that modern H. sapiens arose
from a single ardlaic group in Africa. According to this "Out of
Africa" hypothesis ( also called the replacement hypothesis) the
Neanderthals and other archaic peoples outside Africa were evolutionary dead
ends (Figure 17.44b). Proponents of this hypothesis argue that modern humans
spread out of Africa about 100,000 years ago and completely replaced the archaic
H. sapiens in other regions. So far, genetic evidence mostly; supports the
replacement hypothesis. Based on comparisons of mitochondrial DNA (mtDNA) and
of Y chromosomes between samples from various human populations, today's global
human population seems to be very genetically uniform. Supporters of the
replacement hypothesis argue that such uniformity could only stem from a single
ancestral stock, not from diverse, geographically isolated populations.
Proponents of the multiregional model counter that interbreeding between
populations can explain our great genetic similarity. Debate about which
hypothesis is correct-a multiregional or an "Out of Africa" origin of
modem humans--centers mainly on differences in interpreting the fossil record
and molecular data. As research continues, perhaps anthropologists will come
closer to a consensus on our origins.

Figure 17.44 "Cut of Africa"-but when? Two hypotheses
for the origin of modern Homo
sapiens. There is no question that Africa is the cradle of
humanity; we can all trace our ancestry back to that continent. But how
recently was the ancestor common to all the world's modern humans still in
Africa? (a) According to the multiregional hypothesis, modern humans throughout
the world evolved from the Homo erectus populations that spread to different
regions beginning almost 2 million years ago. Interbreeding between populations
(dashed line in the diagram) kept humans very similar genetically. (b) In
contrast, the "Out of Africa" hypothesis postulates that all regional
descendants of H. erectus became extinct except in Africa. And according to
this model, it was a second dispersal out of Africa, just 100,000 years ago,
that populated the world with modern humans, which had evolved from H. erectus
in Africa.
Cultural Evolution
An erect stance was the most radical anatomical change in our evolution; it
required major remodeling of the foot, pelvis, and vertebral column.
Enlargement of the brain was a secondary alteration made possible by prolonging
the growth period of the skull and its contents (see Figure 14.17). The primate
brain continues to grow after birth, and the period of growth is longer for a
human than for any other primate. The extended period of human development also
lengthens the time parents care for their offspring, which contributes to the
child's ability to benefit from the experiences of earlier generations. This is
the basis of culture—the transmission of accumulated knowledge over
generations. The major means of this transmission is language, written and
spoken.
Cultural
evolution is continuous, but there have been three major stages. The first
stage began with nomads who hunted and gathered food on the African grasslands
2 million years ago. They made tools, organized communal activities, and divided
labor. Beautiful ancient art is just one example of our cultural roots in these
early societies (Figure 17.45) .The second main stage of cultural evolution
came with the development of agriculture in Africa, Eurasia, and the Americas
about 10,000 to 15,000 years ago. Along with agriculture came permanent
settlements and the first cities. The third major stage in our cultural
evolution was the Industrial Revolution, which began in the eighteenth century.
Since then, new technology has escalated exponentially; a single generation
spanned the flight of the Wright brothers and Neil Armstrong's walk on the
moon. It took less than a decade for the World-Wide Web to transform commerce,
communication, and education. Through all this cultural evolution, from simple
hunter-gatherers to high-tech societies, we have not changed biologically in
any significant way. We are probably no more intelligent than our cave-dwelling
ancestors. The same toolmaker who chipped away at stones now designs microchips
and software. The know-how to build. skyscrapers, computers, and spaceships is
stored not in our genes but in the cumulative product of hundreds of
generations of human experience, passed along by parents, teachers, books, and
electronic media.


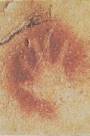


European Bison Rhinocerous Human hand Horses Owl
Figure 17.45 Art History
goes way back, and so does our fascination with and dependence on animal
diversity. Cro-Magnon wildlife artists
created these remarkable paintings beginning
about 30,000 years ago. Three cave explorers found this prehistoric art
gallery on Christmas eve 1994, when they ventured into a cavern near Vllon
–Pont d’Arc in southern France.


Summary
Early Animals and the Cambrian Explosion
Animals probably evolved from a colonial, flagellated protist more than 700
million years ago. At the beginning of the Cambrian period, 545 million years
ago, animal diversity exploded.
.Animal Phylogeny Major branches of animal evolution are
defined by four key evolutionary differences: the presence or absence of true
tissues; radial versus bilateral body symmetry; presence or absence of a body
cavity at least partly lined by mesoderm; and details of embryonic development.
.Sponges Sponges (phylum Porifera)
are sessile animals with porous bodies and choanocytes but no true tissues.
They filter-feed by drawing water through pores in the sides of the body.
.Cnidarians
Cnidarians (phylum Cnidaria) have radial symmetry, a gas-trovascular cavity,
and tentacles with cnidocytes. The body is either a sessile polyp or floating
medusa.
.Flatworms Flatworms (phylum Platyhelminthes) are the
simplest bilatera! animals. They may be free-living or parasitic in or on
plants and animals.
.Roundworms Roundworms (phylum Nematoda) are unsegmented
and cylindrical with tapered ends. They have a complete digestive tract and a
pseudocoelom. They may be free-living or parasitic in plants and animals.
.Mollusks
Mollusks (phylum Mollusca) are soft-bodied animals often protected by a hard
shell. The body has three main parts: a muscular foot, a visceral mass, and a
fold of tissue called the mantle-
.Annelids
Annelids (phylum Annelida) are segmented worms. They may be free-living or
parasitic on other animals.
.Arthropods Arthropods
(phylum Arthropoda) are segmented animals with an exoskeleton and specialized,
jointed appendages. Arthropods consist of four main groups: Arachnids,
crustaceans, millipedes and centipedes, and insects. In species diversity,
insects outnumber all other forms of life combined.
.Echinoderms
Echinoderms (phylum Echinodermata) are sessile or slow-moving marine animals
that lack body segments and possess a unique water vascular system. Bilaterally
symmetrical larvae usually change to radially symmetrical adults. .
.Characteristics of Chordates Chordates (phylum Chordata)
are defined by a dorsal, hollow nerve cord; a flexible notochord; pharyngeal
slits; and a post-anal tail. Tunicates and lancelets are invertebrate
chordates. All other chordates are vertebrates, possessing a cranium and
backbone.
.Fishes Agnathans are jawless vertebrates. Cartilaginous
fishes ( class Chondrichthyes), such as sharks, are mostly predators with
powerful jaws and a flexible skeleton made of cartilage. Bony fishes ( class
Osteichthyes) have a stiff skeleton reinforced by hard calcium salts. Bony
fishes are further classified into ray-finned fishes, lungfishes, and
lobe-finned fishes-
.Amphibians Amphibians ( class Amphibia) are tetrapod
vertebrates that usually deposit their eggs (lacking shells) in water. Aquatic
larvae typically undergo a radical metamorphosis into the adult stage. Their
moist skin requires that amphibians spend much of their adult life in moist
environments.
.Reptiles
Reptiles (class Reptilia) are terrestrial ectotherms with lungs and waterproof
skin covered by scales. Their amniotic eggs enhanced reproduction on land.
.Birds Birds ( class Aves )
are endothermic vertebrates with amniotic eggs, wings, feathers, and other
adaptations for flight.
.Mammals Mammals ( class Mammalia) are endothermic
vertebrates with hair and mammary glands. There are three major groups:
Monotremes lay eggs; marsupials use a placenta but give birth to tiny offspring
that usually complete development while attached to nipples inside the mother's
pouch; and eutherians, or placental mammals, use their placenta in a
longer-lasting association between the mother and her developing young.

Homo Sapiens (modern humans) originated in
Africa about 250,000 and emerged from Africa about 100,000. By 30,000 they are in Siberia, by 14,000 in
Americas and by 1,600 in remote Pacific islands.
Earth’s
New Crisis
Of
the many crises in the history of life, the impact of one species, Homo
sapiens, is the latest and potentially the most devastating.
Evolution of the
human brain may have been anatomically simpler than acquiring an upright
stance, but the global consequences of cerebral expansion have been enormous.
Cultural evolution made Homo sapiens a new force in the history of life-a
species that could defy its physical limitations and shortcut biological
evolution. We do not have to wait to adapt to an environment through natural
selection; we simply change the environment to meet our needs. We are the most
numerous and widespread of all large animals, and everywhere we go, we bring
environmental change. There is nothing new about environmental change. The
history of life is the story of biological evolution on a changing planet. But
it is unlikely that change has ever been as rapid as in the age of humans. Cultural
evolution outpaces biological evolution by orders of magnitude. We are changing
the world faster than many species can adapt; the rate of extinctions in the
twentieth century was 50 times greater than the average for the past 100,000
years.
This rapid rate
of extinction is mainly a result of habitat destruction, which is a function of
human cultural changes and overpopulation. Feeding, clothing, and housing 6
billion people imposes an enormous strain on Earth's capacity to sustain life.
If all these people suddenly assumed the high standard of living enjoyed by
many people in developed nations, It is likely that Earths support systems
would be overwhelmed. Already, for example, current rates of fossil-fuel
consumption, mainly by developed nations, are so great that waste carbon
dioxide may be causing the temperature of the atmosphere to increase enough to
alter world climates. Today, it is not just individual species that are
endangered, but entire ecosystems, the global atmosphere, and the oceans. Tropical
rain forests, which playa vital role in moderating global weather, are being
cut down at a startling rate. Scientists have hardly begun to study these
ecosystems, and many species in them may become extinct before they are even
discovered.